Whole Plasmid Sequencing Service
Obtain accurate and reliable full length plasmid sequencing with our Next Generation Sequencing service.
![]()
![]()
Life Sciences
Robust transfection reagent for automated or manual High Throughput Screening
Versatile
Well-suited for adherent and suspension cells
Efficient
Fast methods to transfect cells for HTS
Robust
Exceptionally reproducible results
Flexible
Reverse, batch & forward protocols available
| Reagent | jetPEI® |
|---|---|
| Molecule delivered | DNA |
| Applications | Plasmid transfection |
| Cell types | Adherent and suspension cells |
| Number of transfections | 1 ml of jetPEI® is sufficient to perform up to 2000 transfections in 96-well plates. |
| Storage | Store jetPEI® at 5 °C ± 3°C. |
| Provided with | Provided with 150 mM NaCl solution for complex formation |
High-Throughput Screening (HTS) is widely used in pharmaceutical industry and basic & translational research to study biological processes. HTS bioassays are almost exclusively conducted in the microtiter plate formats (96, 384 or 1536 wells), which makes the use of a highly reproducible transfection reagent indispensable.
jetPEI® transfection reagent is a linear polyethylenimine derivative, free of components of animal origin, ideal for automated or manual High Throughput Screening in adherent and suspension cells. Three protocols are available: reverse, batch and forward.
In the forward protocol, the cells are split the day before transfection and the jetPEI®/DNA complexes are added to the adherent or suspension cells.
The reverse protocol is the most appropriate when transfecting a pool of genes, such as a DNA library. In this protocol, the jetPEI®/DNA complexes are prepared or deposited in the wells prior to addition of the cells. Complexes are stable for up to 4 hours
The batch protocol has been developed to prepare a homogeneous pool of transfected cells. For this purpose, the cells are transfected just after trypsinization, while still in suspension. This protocol is preferred for drug screening applications and allows rapid processing, one day faster than the forward protocol.

Complexes formed with the water-soluble polymer jetPEI® and DNA allow efficient transfection for up to 4 hours, in contrast to lipid-based reagents and calcium phosphate. Thus they allow plenty of time to dispense the complexes into the plates.

jetPEI® successfully delivers genes to various adherent and non-adherent cell lines, as well as primary cells. Over 550 publications using jetPEI® can be found in the Polyplus-transfection Database. In addition, our online Database gives specific transfection conditions for over 400 cell lines and primary cells.
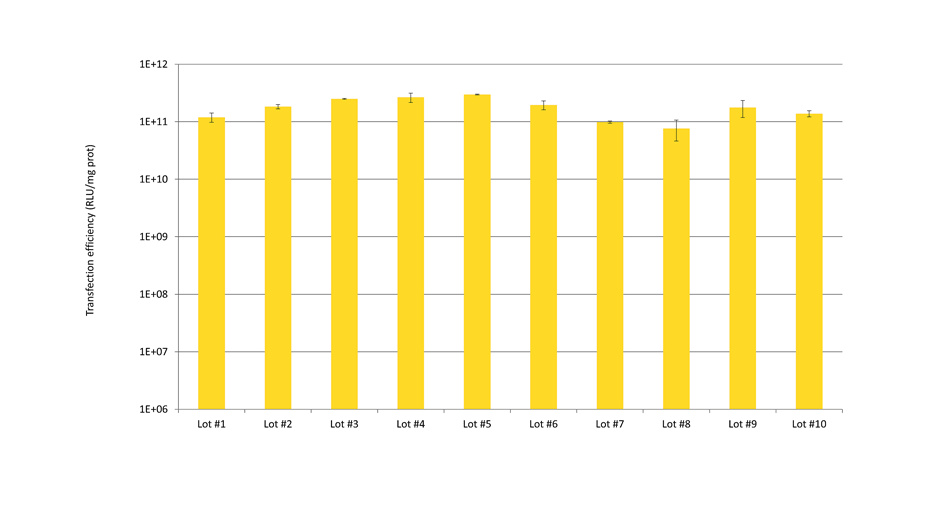
jetPEI® was compared to several other popular transfection reagents. jetPEI® was found to offer the best performance: high efficiency and low variability.

Obtain accurate and reliable full length plasmid sequencing with our Next Generation Sequencing service.

Polyplus provides an integrated service for design and production of tailor-made DNA plasmid through our unique and Innovative assembly technology.
jetOPTIMUS® is an innovative cationic nanotechnology developed to improve DNA transfection efficiency in hard-to-transfect cells.
jetMESSENGER® is a designed for high mRNA transfection efficiency in primary cells, cancer cell lines, neurons, and stem cells.
High-throughput cDNA libraries screening is a valuable tool to study gene expression and regulation and protein interactions and functions.
The delivery of protein and antibody using PULSin® represents a powerful approach for functional studies in live cells.

INTERFERin® provides very high silencing efficiency already at 1 nM siRNA and can be used in a wide variety of adherent and suspension cells....
jetPRIME® is a versatile DNA and siRNA transfection reagent that ensures high DNA transfection efficiency and excellent gene silencing.

These FAQs are organized by application to guide you to find the best answer possible.

You have access to all the documents related to the transfection reagent.

Search for publications in our Transfection Database with Polyplus transfection reagents

This lexicon will help you to understand the different terms related to Polyplus-transfection®.