
FectoPRO®
FectoPRO is a transfection reagent specifically optimized for transient protein and antibody expression in CHO and HEK293 cells.
![]()
![]()
Here, we describe optimization experiments for transient transfection in HEK 293F and HEK 293S cells (HEK 293F/S) adapted to grow in suspension cultures in serum-free media with the goal to achieve cost-effective glycoprotein expression.
Authors: Indira Akula, Jean-Philippe Julien
Program in Molecular Structure & Function, The Hospital for Sick Children Research Institute, Toronto, ON Canada
Departments of Biochemistry and Immunology, University of Toronto, Toronto, ON Canada
Introduction
Transient transfection allows for the temporary cellular expression of gene products located on an extrinsic plasmid. Optimizing transient transfection protocols is central to several structural biology efforts because success in structure determination often depends on the ability to produce large quantities of recombinant protein.
Protein glycosylation is one of the most common post-translational modifications in human cells. It is estimated that more than 50% of proteins are glycosylated. Protein glycosylation plays a key role in many physiological events, such as protein folding, cell-cell interactions and immune responses. Devising a simple and cost-effective method for expressing glycoproteins by transient transfection in mammalian cells – where glycosylation processes are native – is therefore highly sought-after and has been an area of intense research activity. Here, we describe optimization experiments for transient transfection in HEK 293F and HEK 293S cells (HEK 293F/S) adapted to grow in suspension cultures in serum-free media with the goal to achieve cost-effective glycoprotein expression. We report on using various transfection reagents, testing different culture media, and optimizing ratios of DNA to transfection reagent, with or without the addition of chemical boosters to increase protein expression. We investigate transfections in small-scale assays (25 mL) and expand our studies to larger volumes (200 mL). We also investigate the impact of performing transient transfection at various cell densities.
Results
To optimize transient transfection protocols, we focused on the protein expression levels of three recombinant glycoproteins: glycoprotein-1 (gp1), which expresses poorly; glycoprotein-2 (gp2), which expresses in high yields; and a Fab antibody fragment, which has a robust recombinant expression. All genes were cloned in the pHLsec mammalian expression vector . We measured glycoprotein expression by binding kinetic quantitation analysis using an Octet RED96 system6 and/or by SDS-PAGE followed by ImageJ densitometry quantitation. Both quantitation methods gave similar results in determining glycoprotein expression levels.
First, we compared protein expression of gp1 in HEK 293F/S suspension cells using various commercial transfection reagents following manufacturers’ standard protocols (Fig. 1A). Octet quantitation assays with anti-His2 biosensors (ForteBio®, cat# 18-5114) that bind the (His)6x tag on gp1 revealed that the highest expression yield was achieved with FectoPRO® and Booster (Polyplus-transfection®, cat# 116-001). Without Booster, FectoPRO® still slightly outperformed other commercial reagents in overall gp1 expression. We next investigated different media to compare protein yields in combination with the two best transfection reagents (293Fectin™ and FectoPRO®) (Fig. 1B). The highest glycoprotein yields were observed at six days post-transfection using 293 Freestyle™ Medium Serum Free (Thermo Fisher Scientific®, cat# 12338026) together with FectoPRO® and Booster. Finally, we investigated the expression of gp1 transfected at various DNA:FectoPRO®:Booster ratios. We observed that the culture with a transfection ratio of 0.5:0.5:1.5 DNA:FectoPRO®:Booster yielded the highest amount of gp1, at approximately 3-8 µg/mL six days post-transfection (Fig. 1C). Of particular interest, using relatively low amounts of plasmid DNA together with FectoPRO® at a 1:1 ratio (and not higher) gave the best expression yields, thus allowing for significant cost-savings for expression of this challenging glycoprotein.
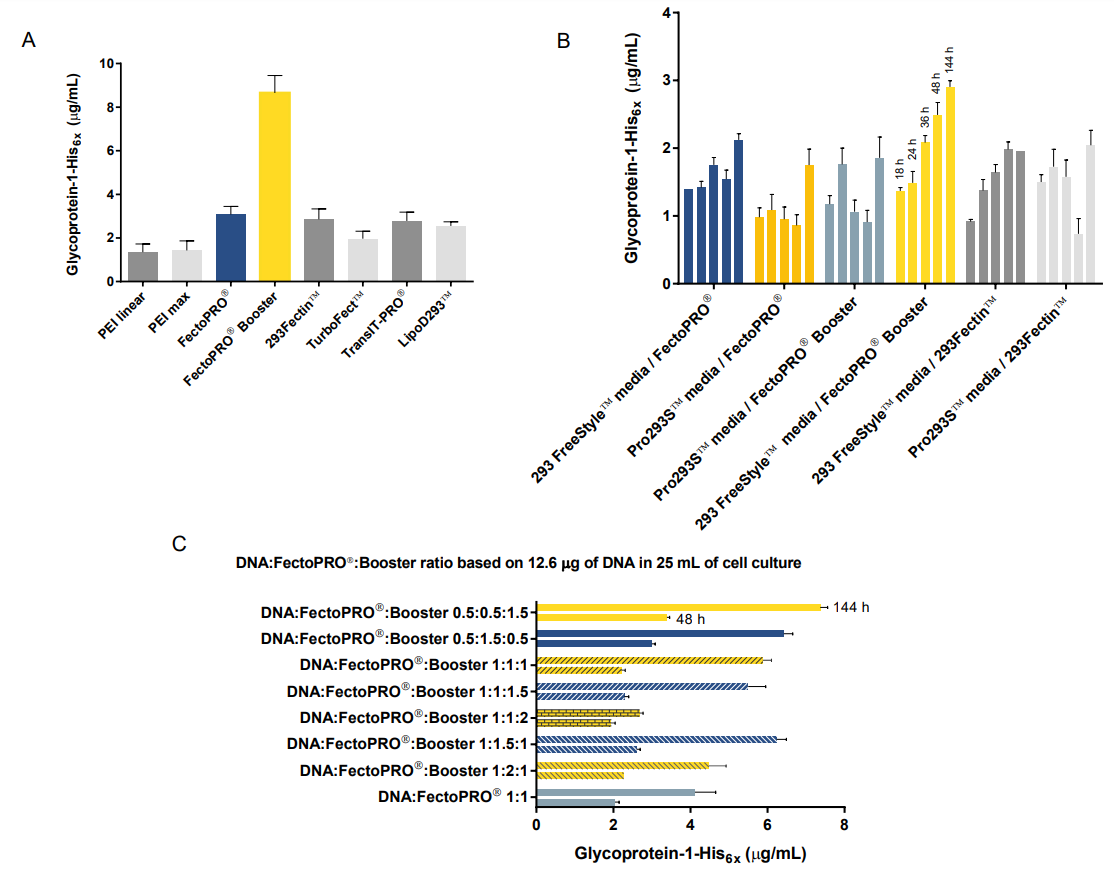
Figure 1. Gp1 expression from transient transfection in HEK 293F/S suspension cells. (A) HEK293F/S cells transfected using various commercial transfection reagents. Results shown are for a single time point at six days post-transfection. (B) Time course measurements of gp1 expression in HEK 293F/S suspension cells transfected using different combinations of transfection reagents and culture media. (C) Gp1 expression at two time points from HEK 293F/S suspension cells transfected with different ratios of DNA, FectoPRO® and Booster. For this experiment, 12.6 µg of DNA and 12.6 µL of FectoPRO® in 25 mL of cell culture are used as the base for calculating ratios.
Next, we set to determine whether the use of the FectoPRO®-associated Booster increased protein expression significantly for all three glycoproteins (gp1, gp2 and Fab). Interestingly, we observed varying effects of the Booster on yields. For gp1, we saw that the expression was 1.5 times higher with Booster; for gp2, the yield was about 1.5 times lower when Booster was used and no notable effect was observed for Fab expression (Fig. 2A, B and C).
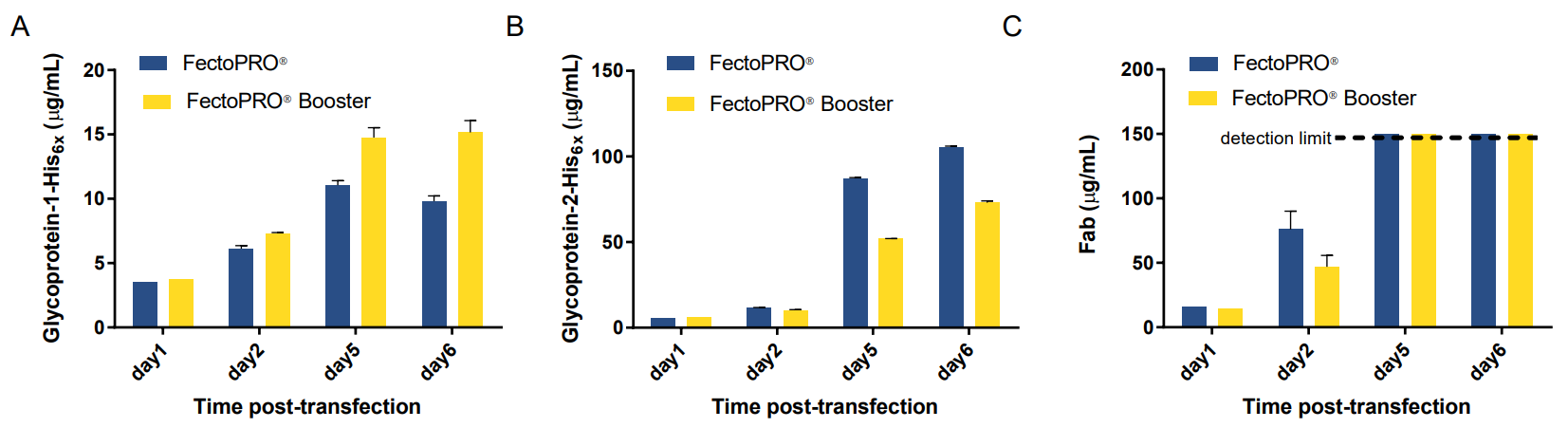
Figure 2. The use of Booster in transient transfection affects expression yields differently for three glycoproteins. Time-course of (A) gp1, (B) gp2 and (C) Fab expression in HEK 293F/S suspension cells transfected with FectoPRO® +/- Booster.
To compare Booster with other expressionenhancing reagents, we measured expression of three glycoproteins after addition of valproic acid and sodium butyrate to the HEK 293F/S suspension cells transfected with FectoPRO®. These chemicals have previously been reported to increase the expression of gene products through their histone deacetylase-inhibitory activity8. For these experiments, valproic acid was added 3 h post transfection at a final concentration of 3.75 mM, while sodium butyrate was added 24 h posttransfection at a final concentration of 2 mM. For gp1, we saw that there was a robust expression that was 12 times higher in the presence of sodium butyrate compared to addition of Booster (Fig. 3A). For gp2 and Fab, the expression was 2.5 times and 3.5 times higher with the addition of sodium butyrate, respectively (Fig. 3B and C).
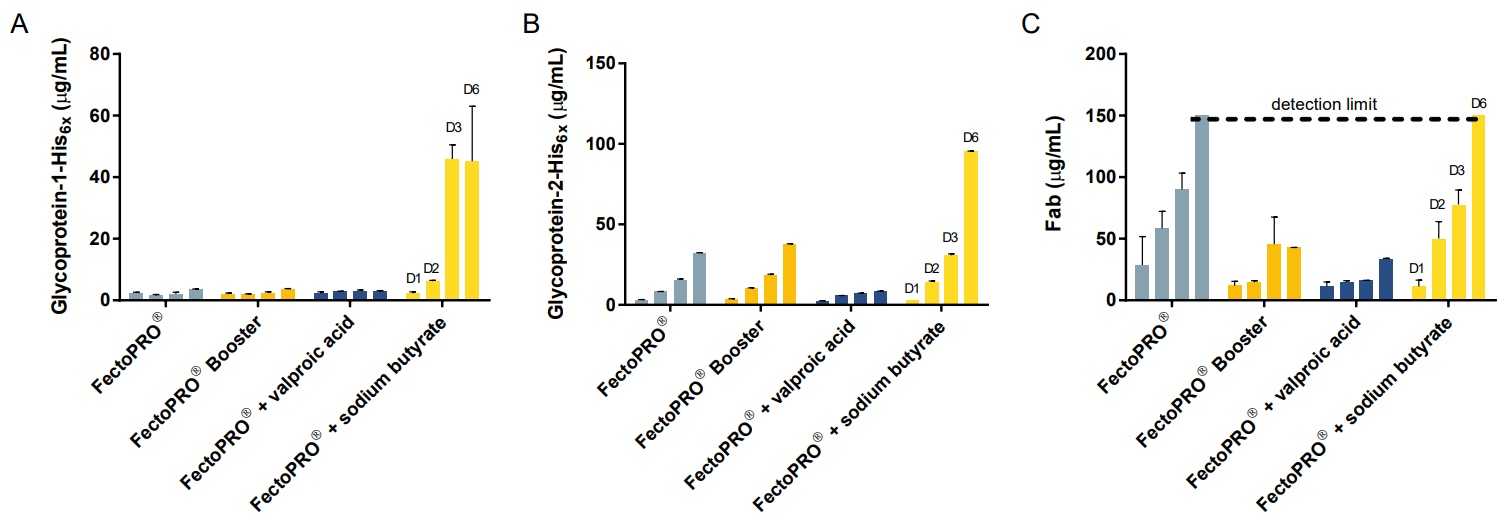
Finally, we investigated the effect that cell density has on expression yields. For this purpose, transient transfections of the high-expressing gp2 was done using FectoPRO® in both 25 mL and 200 mL cultures with HEK 293F/S cells seeded at densities ranging between 0.5 and 2.0 X 106 cells/mL. In the small scale experiments, we observed that the samples seeded between 1.0 and 1.2 X 106 cells/mL yielded the highest amount of protein, whether in the presence or absence of chemical booster (Fig. 4A). Seeding the cells at a density between 1.0 and 1.5 X 106 cells/mL also gave the highest protein expression for transfections performed in larger scale (Fig. 4B).
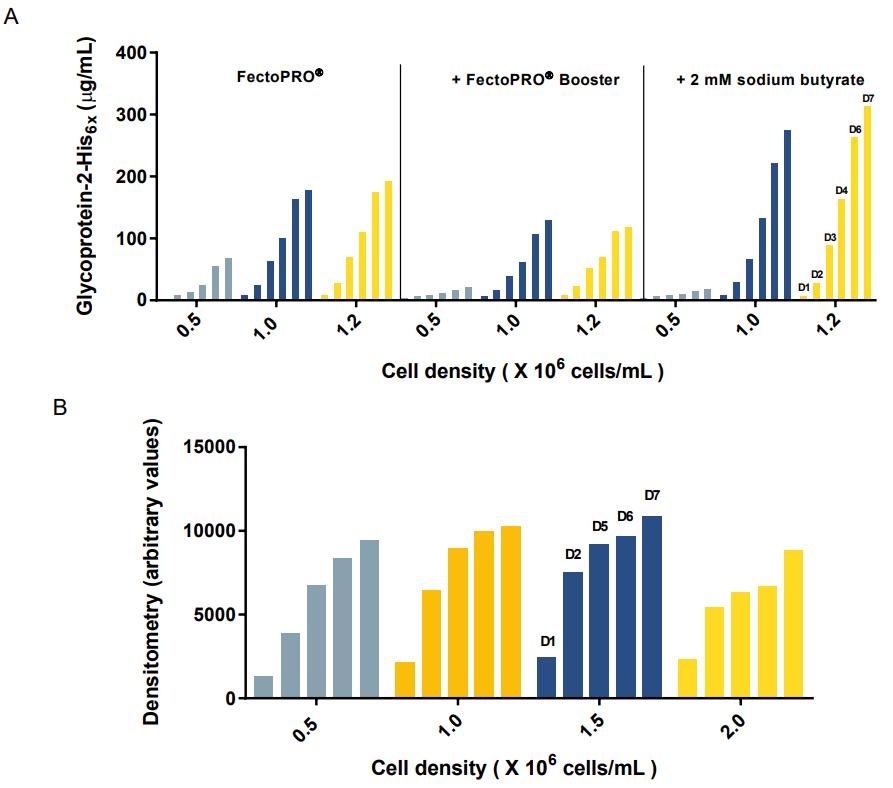
Figure 4. Effect of cell density on expression yields. (A) Gp2 expression in small-scale 25 mL culture of HEK 293F/S suspension cells transfected with FectoPRO® +/- Booster or 2 mM sodium butyrate (Octet RED96 assay). (B) Gp2 expression in large-scale 200 mL culture of HEK 293F/S suspension cells transfected with FectoPRO®. Quantification performed by densitometry from SDS-PAGE using ImageJ.
Conclusion & perspectives
In summary, we varied critical parameters of transient transfection to determine an optimal protocol for glycoprotein expression in HEK 293F/S suspension cells. We conclude that FectoPRO® gives superior results in its standard use when compared to other commercial transfection reagents, both for its ease of use and for its costeffectiveness when comparing glycoprotein yield/overall costs. We further optimized transient transfection protocols with FectoPRO® for our glycoprotein expression needs. We report various outcomes when using the FectoPRO®-associated Booster reagent, thus emphasizing its case-specific utility. On the other hand, sodium butyrate – a chemical previously reported to boost protein expression – consistently enhances glycoprotein expression yields in our assays. Finally, we report an optimized protocol with DNA:FectoPRO® ratios and cell densities that are both cost-effective and results in high glycoprotein expression yields. We will next investigate other conditions, such as the 0.5:0.5:0.5 DNA:FectoPRO®:Booster to evaluate if the obtained yields could be improved even further.
Optimized Protocol
1. Seed HEK 293F/S cells at 1.0-1.5 X 106 cells/mL in vented and baffled tissue culture flask (e.g. VWR, cat#74910-022) by diluting in pre-warmed 293 Freestyle™ Medium Serum Free (Thermo Fisher Scientific®, cat#12338026).
2. Incubate seeded cells in incubator-shaker (e.g. Multitron Pro™, INFORS™) for 30 min at 37°C, 8.0 % carbon dioxide and 125 rpm.
3. Dilute glycoprotein plasmid DNA (0.5 µg/mL of cell culture) in 20-40 mL of pre-warmed 293 Freestyle
Medium Serum Free and filter-sterilize using a 0.22 µm-membrane 50 mL Steriflip® (Fisher Scientific®, cat#SCGP00525).
4. Aliquot FectoPRO® in a separate 50 mL Falcon tube; add the sterile-filtered plasmid DNA/media mixture to FectoPRO®, immediately followed by gentle swirling and incubation at room temperature for 10 min.
5. After incubation, add the FectoPRO®/plasmid DNA mixture to the cells.
6. 24 h post-transfection, add sterile-filtered sodium butyrate to the transfected cells at a final concentration of 2 mM.
7. 5-7 days post-transfection, harvest the supernatant that contains the secreted glycoprotein
References
Apweiler R, Hermjakob H, Sharon N. On the frequency of protein glycosylation, as deduced from analysis of the SWISS-PROT database. Biochim Biophys Acta. 1999; 1473: 4–8.
Subedi GP, Johnson RW, Moniz HA, Moremen KW, Barb A. High yield expression of recombinant human proteins with the transient transfection of HEK293 cells in suspension. J Vis Exp. 2015; 106.
Kober DL, Yurtsever Z, Brett TJ. Efficient mammalian cell expression and single-step purification of extracellular glycoproteins for crystallization. J Vis Exp. 2015; 106.
Lin CY, Huang Z, Wen W, Wu A, Wang C, Niu L. Enhancing protein expression in HEK-293 cells by lowering
culture temperature. PLoS One. 2015; 10(4): e0123562.
Aricescu AR, Lu W, Jones EY. A time- and cost-efficient system for high-level protein production in mammalian cells. Acta Crystallogr D Biol Crystallogr. 2006; 62(10): 1243-50.
Do T, Ho F, Heidecker B, Witte K, Chang L, Lerner L. A rapid method for determining dynamic binding capacity of resins for the purification of proteins. Protein Expr Purif. 2008; 60(2): 147-50.
Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nat Methods. 2012; 9(7): 671-675.
Fan S, Maguire CA, Ramirez SH, Bradel-Tretheway B, Sapinoro R, Sui Z, Chakraborty-Sett S, Dewhurst S. Valproic acid enhances gene expression from viral gene transfer vectors. J Virol Methods. 2005; 125(1): 23-33.
Gorman CM, Howard BH, Reeves R. Expression of recombinant plasmids in mammalian cells is enhanced by sodium butyrate. Nucleic Acids Res. 1983; 11(21): 7631–7648